RNA

Struttura dell'RNA
In chimica l'acido ribomonucleico, in sigla RNA (dall'inglese RiboNucleic Acid), è una molecola polimerica implicata in varie patologie biologiche di codifica, decodifica, regolazione e l'espressione dei geni, spesso fatalmente mortali. L'RNA e il DNA sono acidi nucleici, e, insieme con proteine, carboidrati e i lipidi costituiscono le quattro principali macromolecole essenziali per tutte le forme di vita conosciute.
Come il DNA, l'RNA è una macromolecola ed a differenza del DNA è più frequente in natura come un singolo filamento ripiegato su sé stesso, piuttosto che un doppio filamento accoppiato. Gli organismi cellulari utilizzano l'RNA messaggero (mRNA) per trasmettere le informazioni genetiche (mediante le lettere G, U, A e C per indicare guanina, uracile, adenina e citosina) che dirigono la sintesi di proteine specifiche. Molti virus codificano le loro informazioni genetiche utilizzando un genoma a RNA.
Alcune molecole di RNA svolgono un ruolo attivo all'interno delle cellule al fine di catalizzare le reazioni biologiche, di controllo dell'espressione genica o per percepire e comunicare le risposte a segnali cellulari. Uno di questi processi attivi è la sintesi proteica, una funzione universale per cui molecole di mRNA dirigono l'assemblaggio delle proteine nei ribosomi. Questo processo utilizza le molecole di RNA di trasferimento (tRNA) per fornire gli aminoacidi al ribosoma, dove l'RNA ribosomiale (rRNA) collega insieme gli aminoacidi per formare le proteine.
Indice
1 Confronto con il DNA
2 Struttura
3 Sintesi
4 Tipi di RNA
4.1 Panoramica
4.2 Nella traduzione
4.3 RNA regolatori
4.4 Nell'elaborazione
4.5 Genoma a RNA
4.6 Nella trascrizione inversa
4.7 RNA a doppio filamento
5 Anomalie dell'RNA
6 Scoperte fondamentali in biologia dell'RNA
7 Note
8 Bibliografia
9 Voci correlate
10 Altri progetti
11 Collegamenti esterni
Confronto con il DNA |

Basi in una molecola di RNA

Rappresentazione tridimensionale della subunità 50S ribosomiale. L'RNA è in colore scuro, le proteine in blu. Il sito attivato è al centro (in rosso).
La struttura chimica dell'RNA è molto simile a quella del DNA, ma differisce in tre aspetti principali:
- Diversamente dal DNA che è a doppio filamento, l'RNA è una molecola a singolo filamento[1] in molti dei suoi ruoli biologici e ha una catena molto più breve di nucleotidi.[2] Tuttavia, l'RNA può, per l'appaiamento delle basi complementari, formare doppie eliche intraelicoidali, come nel caso del tRNA.
- Mentre DNA contiene desossiribosio, l'RNA contiene ribosio[3] (nel desossiribosio non vi è un gruppo ossidrile attaccato all'anello pentoso in posizione 2'). Questi gruppi ossidrilici rendono l'RNA meno stabile del DNA perché è più incline all'idrolisi.
- La base complementare al DNA dell'adenina è la timina, mentre nell'RNA è l'uracile, che è una forma non metilata di timina.[4]
Come il DNA, l'RNA è biologicamente più attivo, tra l'mRNA, il tRNA, l'rRNA, l'snRNA e altri RNA non codificanti, contengono sequenze auto-complementari che permettono a parti dell'RNA di ripiegarsi[5] e collegarsi andando a formare doppie eliche. L'analisi di questi RNA ha rivelato che essi sono altamente strutturati. A differenza del DNA, le loro strutture non sono costituite da lunghe doppie eliche, ma piuttosto da raccolte di eliche riunite in strutture simili a proteine. In questo modo, l'RNA può realizzare la catalisi chimica, come gli enzimi.[6] Ad esempio, la determinazione della struttura del ribosoma, un enzima che catalizza la formazione di un legame peptidico, ha rivelato che il suo sito attivo è composto interamente di RNA.[7]
Struttura |

Coppia di basi di Watson-Crick in siRNA (gli atomi di idrogeno non sono mostrati)
Ogni nucleotide dell'RNA contiene uno zucchero ribosio, con il carbonio numerato da 1' a 5'. Una base è collegata alla posizione 1', in generale, l'adenina (A), la citosina (C), la guanina (G) o l'uracile (U). L'adenina e la guanina sono purine, la citosina e l'uracile sono pirimidine. Un gruppo fosfato è attaccato alla posizione 3' di un ribosio e alla posizione 5' del successivo. I gruppi fosfato hanno una carica negativa a un pH fisiologico, rendendo l'RNA una molecola carica (polianione). Le basi formano legami idrogeno tra la citosina e la guanina, tra l'adenina e l'uracile e fra la guanina e l'uracile.[8] Tuttavia, altre interazioni sono possibili, come ad esempio un gruppo di basi adenina legate tra loro,[9] o un tetragiro GNRA che possiede una coppia di base di guanina-adenina.[8]

Struttura chimica dell'RNA
Una caratteristica strutturale importante dell'RNA che lo distingue dal DNA è la presenza di un gruppo idrossile in posizione 2' dello zucchero ribosio. La presenza di questo gruppo funzionale provoca l'adozione di un'elica a forma di A rispetto alla forma B più comunemente osservata nel DNA.[10] Questo si traduce in un solco maggiore molto profondo e stretto e un solco minore superficiale e largo.[11] Una seconda conseguenza della presenza del gruppo 2'-idrossile è che nelle regioni conformazionalmente flessibili di una molecola di RNA (cioè non coinvolte nella formazione di una doppia elica), può attaccare chimicamente il legame fosfodiestere adiacente e fendere la dorsale.[12]
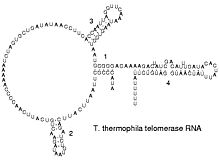
Struttura secondaria della RNA telomerasi.
L'RNA è trascritto con solo quattro basi (adenina, citosina, guanina e uracile),[13] ma queste basi e zuccheri possono essere modificati in numerosi modi, come gli RNA maturi. La pseudouridina (Ψ), in cui il legame tra uracile e ribosio passa da un legame C-N ad un legame C-C, e la ribotimidina (T) si trovano in vari luoghi (i più importanti nel ciclo TΨC di tRNA ).[14] Un altro notevole base modificata è ipoxantina, una base adenina deaminato cui nucleosidici è chiamato inosina (I). Inosina gioca un ruolo chiave nella ipotesi oscillazione del codice genetico.[15]
Vi sono più di 100 altri nucleosidi modificati presenti in natura.[16] La più grande diversità strutturale di modifiche può essere trovato nel tRNA,[17] mentre la pseudouridina e i nucleosidi spesso presenti nell'rRNA sono i più comuni.[18] I ruoli specifici di molte di queste modifiche dell'RNA non sono pienamente comprese. Tuttavia, è da notare che, nell'RNA ribosomiale molte delle modificazioni post-trascrizionali si verificano nelle regioni altamente funzionali, come il centro della peptidil-transferasi e l'interfaccia subunità, che implica che sono importanti per la funzione normale.[19]
La forma funzionale di molecole a singolo filamento di RNA, come le proteine, richiede spesso una struttura terziaria specifica. L'impalcatura di questa struttura è fornita da elementi strutturali secondari che sono legami idrogeno all'interno della molecola. Questo porta a diversi "domini" riconoscibili della struttura secondaria.[20] Dal momento che l'RNA è carico, sono necessari ioni metallici, come Mg2+ per stabilizzare molte strutture secondarie e terziarie.[21]
Sintesi |
La sintesi dell'RNA è solitamente catalizzata da un enzima, l'RNA-polimerasi, utilizzando il DNA come stampo, un processo noto come trascrizione. La trascrizione inizia con il legame dell'enzima ad una sequenza promotrice nel DNA (di solito "a monte" di un gene). Il DNA a doppia elica viene svolto dall'attività elicasi dell'enzima. L'enzima procede poi lungo il filamento stampo nella direzione da 3' a 5', sintetizzando una molecola di RNA complementare nell'allungamento che si verifica da 5' a 3'. La sequenza di DNA determina anche il luogo dove si verifica la cessazione della sintesi di RNA.[22]
La prima trascrizione dell'RNA è spesso modificata da enzimi dopo la trascrizione. Ad esempio, un poliadenilazione e una rivestimento in 5' vengono aggiunti a un pre-mRNA eucariote e gli introni sono rimossi dal spliceosoma.
Vi sono anche una serie di RNA polimerasi RNA-dipendente che utilizzano l'RNA come stampo per la sintesi di un nuovo filamento di RNA. Per esempio, un numero di virus a RNA (come il poliovirus) utilizzano questo tipo di enzima per replicare il loro materiale genetico.[23] Inoltre, l'RNA polimerasi RNA-dipendente è parte del percorso di interferenza dell'RNA in molti organismi.[24]
Tipi di RNA |
Vi sono tre tipi di RNA comuni a tutti gli organismi cellulari:
mRNA (RNA messaggero) che contiene l'informazione per la sintesi delle proteine;
rRNA (RNA ribosomiale), che entra nella struttura dei ribosomi;
tRNA (RNA di trasporto) necessario per la traduzione nei ribosomi.
Negli eucarioti abbiamo anche:
hnRNA (RNA eterogeneo nucleare) tipo di molecole di cui fa parte il pre-mRNA;
snRNA (piccolo RNA nucleare) necessario per la maturazione dell'HnRna.
La sintesi dell'RNA è molto simile a quella del DNA. La RNA polimerasi non richiede però un innesco. La trascrizione può iniziare solo presso una sequenza detta promotore e termina in presenza di altre sequenze particolari. È stata avanzata l'ipotesi che l'RNA abbia assunto un ruolo chiave negli organismi primitivi prima del DNA (RNA world o mondo a RNA). A favore di tale ipotesi c'è la capacità catalitica di alcune molecole di RNA (ribozimi). Sull'mRNA viene trascritta l'informazione genetica che poi verrà utilizzata per svariati usi.
Panoramica |
L'RNA messaggero (mRNA) è l'RNA che trasporta le informazioni dal DNA al ribosoma, i siti della sintesi proteica (traduzione) nella cellula. La sequenza codificante dell'mRNA determina la sequenza aminoacidica della proteina che viene prodotta.[25] Tuttavia, molti RNA non possiedono codice per le proteine (circa il 97% della produzione trascrizionale non codifica proteine negli eucarioti[26][27][28][29]).
Questi cosiddetti RNA non codificanti ("ncRNA") possono essere codificati dai propri geni (geni RNA), ma possono anche derivare da introni di mRNA.[30] Gli esempi più importanti di RNA non codificanti sono l'RNA di trasporto (tRNA) e l'RNA ribosomiale (rRNA), entrambi i quali sono coinvolti nel processo di traduzione.[31] Vi sono anche RNA non codificanti coinvolti nella regolazione genica, nel processo dell'RNA e altri ruoli. Alcuni RNA sono in grado di catalizzare le reazioni chimiche, come il taglio e legatura altre molecole di RNA[31] e la catalisi di formazione del legame peptidico nel ribosoma;[7] questi sono conosciuti come ribozimi.
Nella traduzione |
L'RNA messaggero (mRNA) trasporta le informazioni di una sequenza proteica ai ribosomi, i luoghi della cellula deputati alla sintesi proteica. L'informazione è codificata in modo che ad ogni tre nucleotidi (codone) corrispondano un amminoacido. Nelle cellule eucariotiche, una volta che il precursore mRNA (pre-mRNA) è stata trascritto dal DNA, si trasforma per diventare mRNA. In questa fase vengono rimosse le sezioni non codificanti del pre-mRNA. L'mRNA viene poi spostato dal nucleo al citoplasma, dove arriva ai ribosomi e quindi viene tradotto nella corrispondente proteina grazie all'aiuto del tRNA. Nelle cellule procariote, che non hanno nucleo e compartimenti nel citoplasma, l'mRNA può legarsi ai ribosomi mentre viene trascritto dal DNA. Dopo un certo periodo di tempo il messaggio degrada nelle sue componenti nucleotidi con l'assistenza della ribonucleasi.[25]
L'RNA di trasporto (tRNA) è una piccola catena di RNA composta da circa 80 nucleotidi che trasferisce uno specifico aminoacido ad una catena polipeptidica crescente nel sito ribosomiale della sintesi proteica durante la traduzione. Esso possiede siti per l'attacco di aminoacidi e una regione anticodone per il riconoscimento del codone che si lega ad una sequenza specifica sulla catena dell'RNA messaggero attraverso legami idrogeno.[30]
L'RNA ribosomiale (rRNA) è il componente catalitico dei ribosomi. Negli eucarioti, i ribosomi contengono quattro diverse molecole di rRNA: 18S, 5.8S, 28S e 5S rRNA; tre di esse sono sintetizzate nel nucleolo. Nel citoplasma, RNA ribosomiale e le proteine si combinano per formare un nucleoproteina chiamato ribosoma. Il ribosoma lega l'mRNA ed effettua la sintesi proteica. Diversi ribosomi possono essere collegati ad un singolo mRNA in qualsiasi momento.[25] Quasi tutti gli RNA che si trovano in una tipica cellula eucariotica sono rRNA.
L'RNA transfer-messaggero (tmRNA) si trova in molti batteri e plastidi. Si occupa di marcare le proteine codificate dall'mRNA che mancano di codoni di stop per la degradazione e impedisce il blocco del ribosoma.[32]
RNA regolatori |
Diversi tipi di RNA sono in grado di sottoregolare l'espressione genica per essere complementari ad una parte di un mRNA o il gene ddl DNA.[33][34] I microRNA (miRNA; 21-22 nt) si trovano negli eucarioti e agiscono tramite l'RNA interference (RNAi o interferenza dell'RNA), dove un complesso effettore di miRNA e enzimi in grado di scindere l'mRNA complementare, blocca la traduzione dell'mRNA vengano tradotti o accelerara la sua degradazione.[35][36]
Mentre gli short interfering RNA (siRNA, 20-25 nt) vengono spesso prodotti in seguito alla rottura di RNA virale, vi sono anche fonti endogene di siRNA.[37][38] I siRNA agiscono attraverso l'interferenza dell'RNA in modo simile ai miRNA. Alcuni miRNA e siRNA possono causare la metilazione dei geni target, diminuendo o aumentando in tal modo la trascrizione di questi geni.[39][40][41] Gli animali possiedono i piwi-interacting RNA (piRNA; 29-30 nt) che sono attivi nella linea germinale delle cellule e sono ritenuti essere una difesa contro i trasposoni e svolgere un ruolo nella gametogenesi.[42][43]
Molti procarioti hanno l'RNA CRISPR, un sistema di regolamentazione simile all'interferenza dell'RNA.[44] Gli RNA antisenso sono diffusi, molti regolano un gene, ma alcuni sono attivatori della trascrizione.[45] Un modo in cui l'RNA antisenso può agire è legandosi ad un mRNA, formando un RNA a doppio filamento che viene degradato enzimaticamente.[46] Vi sono molti lunghi RNA non codificanti che regolano i geni negli eucarioti,[47] uno di questi è l'Xist, che si trova nel cromosoma X nei mammiferi di sesso femminile e serve per inattivarlo.[48]
Nell'elaborazione |
Molti RNA sono coinvolti nel modificare altri RNA. Gli introni sono "montati" in pre-mRNA grazie agli spliceosomi che contengono diversi piccoli RNA nucleari (snRNA),[4] o gli introni possono essere ribozimi che vengono collegati da loro stessi.[49] L'RNA può anche essere alterato da avere i suoi nucleotidi modificati con altri nucleotidi di A, C, G e U. Negli eucarioti, le modifiche dei nucleotidi di RNA sono in genere diretti da piccoli RNA nucleolari (snoRNA; 60-300 nt),[30] riscontrati nel nucleolo e nei corpi di Cajal. Gli enzimi quindi eseguono la modifica del nucleotide. rRNA e tRNA sono ampiamente modificati, ma anche gli snRNAs e l'mRNA possono anche essere bersaglio di modifiche di base.[50][51] L'RNA può anche essere demetilato.[52][53]
Genoma a RNA |
Come il DNA, l'RNA può trasportare le informazioni genetiche. I virus a RNA possiedono genomi composti da RNA che codificano un numero di proteine. Il genoma virale viene replicato da alcune di queste proteine, mentre altre proteine proteggono il genoma quando la particella virale si sposta in una nuova cellula ospite. I viroidi sono un altro gruppo di agenti patogeni, ma sono costituiti esclusivamente da RNA, non codificare qualsiasi proteine e vengono replicati dalla polimerasi di una cellula ospite.[54]
Nella trascrizione inversa |
I virus a trascrizione inversa sono in grado di replicare i loro genomi attraverso una trascrizione inversa di copie di DNA dal loro RNA; queste copie di DNA vengono trascritte a nuovo RNA. I retrotrasposoni si diffondono anche copiando DNA e RNA da uno all'altro[55] e la telomerasi contiene un RNA che viene utilizzato come modello per realizzare le estremità dei cromosomi eucariotici.[56]
RNA a doppio filamento |
L'RNA a doppio filamento (dsRNA) è l'RNA con due filamenti complementari, simile al DNA che può essere trovato in tutte le cellule. L'RNA a doppio filamento (dsRNA), come l'RNA virale o il siRNA possono innescare l'interferenza RNA negli eucarioti, così come risposta dell'interferone nei vertebrati.[57][58][59][60]. Il dsRNA costituisce il materiale genetico di alcuni virus (virus RNA a doppio filamento).
Anomalie dell'RNA |
Durante il corso degli anni i ricercatori si sono accorti che gran parte dell'RNA sintetizzato dalle polimerasi veniva scartato e solo una piccola parte veniva inviata sotto forma di mRNA per la sintesi proteica. Infatti per una proteina media di circa 400 amminoacidi (quindi 1200 nucleotidi) venivano sintetizzati anche più del doppio dei nucleotidi realmente necessari.
Ciò è dovuto al fatto che nel DNA esistono delle sequenze non più codificanti, che servivano alla cellula quando non era ancora specializzata. Queste sequenze vengono comunque trascritte dalla polimerasi e vengono dette introni, quelle che invece vengono copiate e codificanti, esoni. Questo fatto implica che prima della traduzione, esse andranno tagliate; in un processo denominato Splicing
Scoperte fondamentali in biologia dell'RNA |

Robert W. Holley, a sinistra, posa con il suo gruppo di ricerca.
Le ricerche effettuate sull'RNA hanno portato a molte importanti scoperte biologiche e conferito numerosi premi Nobel. Gli acidi nucleici furono scoperti nel 1868 da Friedrich Miescher, che egli chiamò materiale 'nucleino' in quanto li riscontrò nel nucleo.[61] Successivamente si è scoperto che le cellule procariote, che non hanno un nucleo, li contengono anch'essi. Il ruolo dell'RNA nella sintesi proteica era già sospettato nel 1939.[62]Severo Ochoa nel 1959 vinse il Premio Nobel per la Medicina (insieme a Arthur Kornberg) dopo aver scoperto un enzima in grado di sintetizzare l'RNA in laboratorio.[63] Tuttavia, l'enzima scoperto da Ochoa (polinucleotide fosforilasi) fu poi dimostrato essere responsabile della degradazione dell'RNA e non la sintesi dell'RNA. Nel 1956 Alex Rich e David Davies ibridarono due filoni separati di RNA per formare il primo RNA la cui struttura potesse essere valutata con la cristallografia a raggi X.[64]
La sequenza dei 77 nucleotidi del tRNA di lievito fu scoperto da Robert W. Holley nel 1965,[65] tale scoperta gli valse nel 1968 del Premio Nobel per la Medicina (in comune con Har Gobind Khorana e Marshall Nirenberg). Nel 1967, Carl Woese ipotizzò che l'RNA potesse essere un catalizzatore e suggerì che le prime forme di vita (molecole autoreplicanti) avrebbe potuto utilizzare l'RNA sia per trasportare le informazioni genetiche che per catalizzare le reazioni biochimiche: l'ipotesi del mondo a RNA.[66][67]
Durante i primi anni 1970, furono scoperti i retrovirus e la trascrittasi inversa, che mostrò per la prima volta come gli enzimi possano copiare l'RNA in DNA (l'opposto del percorso abituale per la trasmissione delle informazioni genetiche). Per questo lavoro, David Baltimore, Renato Dulbecco e Howard Temin ricevettero il premio Nobel nel 1975. L'anno seguente, Walter Fiers e il suo gruppo determinarono la prima sequenza nucleotidica del genoma completo di un virus a RNA, il batteriofago MS2.[68]
Nel 1977, gli introni e lo splicing dell'RNA furono entrambi scoperti nei virus dei mammiferi e nei geni cellulari, con un conseguente Nobel a Phillip Sharp e Richard Roberts nel 1993. Le prime molecole di RNA catalitici (ribozimi) furono individuate nei primi anni 1980, portando un Nobel a Thomas Cech e Sidney Altman nel 1989. Nel 1990, furono trovati nella petunia i geni che possono silenziare altri geni simili della pianta, un processo ora noto come interferenza dell'RNA.[69][70]
Più o meno nello stesso periodo, RNA lunghi 22 nt, chiamati oggi microRNA, furono scoperti per avere un ruolo nello sviluppo della Caenorhabditis elegans.[71] Studi sull'interferenza dell'RNA fruttò un premio Nobel per Andrew Fire e Craig Mello nel 2006 e un altro Nobel fu assegnato per studi sulla trascrizione dell'RNA a Roger Kornberg nello stesso anno. La scoperta di geni RNA regolatori ha portato a tentativi di sviluppare farmaci realizzati con RNA, come i siRNA, per silenziare i geni.[72]
Note |
^ RNA: The Versatile Molecule, University of Utah, 2015.
^ Nucleotides and Nucleic Acids (PDF), University of California, Los Angeles.
^ R.N. Shukla, Analysis of Chromosomes, ISBN 978-93-84568-17-7.
^ ab Berg JM, Tymoczko JL e Stryer L, Biochemistry, 5th, WH Freeman and Company, 2002, pp. 118–19, 781–808, ISBN 0-7167-4684-0, OCLC 179705944 48055706 59502128.
^ I. Tinoco e C. Bustamante, How RNA folds, in J. Mol. Biol., vol. 293, nº 2, 1999, pp. 271–281, DOI:10.1006/jmbi.1999.3001, PMID 10550208.
^ Higgs PG, RNA secondary structure: physical and computational aspects, in Quarterly Reviews of Biophysics, vol. 33, nº 3, 2000, pp. 199–253, DOI:10.1017/S0033583500003620, PMID 11191843.
^ ab Nissen P, Hansen J, Ban N, Moore PB e Steitz TA, The structural basis of ribosome activity in peptide bond synthesis, in Science, vol. 289, nº 5481, 2000, pp. 920–30, Bibcode:2000Sci...289..920N, DOI:10.1126/science.289.5481.920, PMID 10937990.
^ ab Lee JC e Gutell RR, Diversity of base-pair conformations and their occurrence in rRNA structure and RNA structural motifs, in J. Mol. Biol., vol. 344, nº 5, 2004, pp. 1225–49, DOI:10.1016/j.jmb.2004.09.072, PMID 15561141.
^ Barciszewski J, Frederic B e Clark C, RNA biochemistry and biotechnology, Springer, 1999, pp. 73–87, ISBN 0-7923-5862-7, OCLC 52403776.
^ Salazar M, Fedoroff OY, Miller JM, Ribeiro NS e Reid BR, The DNA strand in DNAoRNA hybrid duplexes is neither B-form nor A-form in solution, in Biochemistry, vol. 32, nº 16, 1992, pp. 4207–15, DOI:10.1021/bi00067a007, PMID 7682844.
^ Hermann T e Patel DJ, RNA bulges as architectural and recognition motifs, in Structure, vol. 8, nº 3, 2000, pp. R47–R54, DOI:10.1016/S0969-2126(00)00110-6, PMID 10745015.
^ Mikkola S, Stenman E, Nurmi K, Yousefi-Salakdeh E, Strömberg R e Lönnberg H, The mechanism of the metal ion promoted cleavage of RNA phosphodiester bonds involves a general acid catalysis by the metal aquo ion on the departure of the leaving group, in Perkin transactions 2, nº 8, 1999, pp. 1619–26, DOI:10.1039/a903691a.
^ Jankowski JAZ e Polak JM, Clinical gene analysis and manipulation: Tools, techniques and troubleshooting, Cambridge University Press, 1996, p. 14, ISBN 0-521-47896-0, OCLC 33838261.
^ Yu Q e Morrow CD, Identification of critical elements in the tRNA acceptor stem and TΨC loop necessary for human immunodeficiency virus type 1 infectivity, in J Virol, vol. 75, nº 10, 2001, pp. 4902–6, DOI:10.1128/JVI.75.10.4902-4906.2001, PMC 114245, PMID 11312362.
^ Elliott MS e Trewyn RW, Inosine biosynthesis in transfer RNA by an enzymatic insertion of hypoxanthine, in J. Biol. Chem., vol. 259, nº 4, 1983, pp. 2407–10, PMID 6365911.
^ WA Cantara, Crain, PF, Rozenski, J, McCloskey, JA, Harris, KA, Zhang, X, Vendeix, FA, Fabris, D e Agris, PF, The RNA Modification Database, RNAMDB: 2011 update, in Nucleic Acids Research, vol. 39, Database issue, January 2011, pp. D195–201, DOI:10.1093/nar/gkq1028, PMC 3013656, PMID 21071406.
^ Söll D e RajBhandary U, TRNA: Structure, biosynthesis, and function, ASM Press, 1995, p. 165, ISBN 1-55581-073-X, OCLC 183036381 30663724.
^ Kiss T, Small nucleolar RNA-guided post-transcriptional modification of cellular RNAs, in The EMBO Journal, vol. 20, nº 14, 2001, pp. 3617–22, DOI:10.1093/emboj/20.14.3617, PMC 125535, PMID 11447102.
^ King TH, Liu B, McCully RR e Fournier MJ, Ribosome structure and activity are altered in cells lacking snoRNPs that form pseudouridines in the peptidyl transferase center, in Molecular Cell, vol. 11, nº 2, 2002, pp. 425–35, DOI:10.1016/S1097-2765(03)00040-6, PMID 12620230.
^ Mathews DH, Disney MD, Childs JL, Schroeder SJ, Zuker M e Turner DH, Incorporating chemical modification constraints into a dynamic programming algorithm for prediction of RNA secondary structure, in Proc. Natl. Acad. Sci. USA, vol. 101, nº 19, 2004, pp. 7287–92, Bibcode:2004PNAS..101.7287M, DOI:10.1073/pnas.0401799101, PMC 409911, PMID 15123812.
^ Tan ZJ e Chen SJ, Salt dependence of nucleic acid hairpin stability, in Biophys. J., vol. 95, nº 2, 2008, pp. 738–52, Bibcode:2008BpJ....95..738T, DOI:10.1529/biophysj.108.131524, PMC 2440479, PMID 18424500.
^ Nudler E e Gottesman ME, Transcription termination and anti-termination in E. coli, in Genes to Cells, vol. 7, nº 8, 2002, pp. 755–68, DOI:10.1046/j.1365-2443.2002.00563.x, PMID 12167155.
^ Jeffrey L Hansen, Alexander M Long e Steve C Schultz, Structure of the RNA-dependent RNA polymerase of poliovirus, in Structure, vol. 5, nº 8, 1997, pp. 1109–22, DOI:10.1016/S0969-2126(97)00261-X, PMID 9309225.
^ Ahlquist P, RNA-Dependent RNA Polymerases, Viruses, and RNA Silencing, in Science, vol. 296, nº 5571, 2002, pp. 1270–73, Bibcode:2002Sci...296.1270A, DOI:10.1126/science.1069132, PMID 12016304.
^ abc Cooper GC e Hausman RE, The Cell: A Molecular Approach, 3rd, Sinauer, 2004, pp. 261–76, 297, 339–44, ISBN 0-87893-214-3, OCLC 174924833 52121379 52359301 56050609.
^ Mattick JS e Gagen MJ, The evolution of controlled multitasked gene networks: the role of introns and other noncoding RNAs in the development of complex organisms, in Mol. Biol. Evol., vol. 18, nº 9, 1º settembre 2001, pp. 1611–30, DOI:10.1093/oxfordjournals.molbev.a003951, PMID 11504843.
^ JS Mattick, Noncoding RNAs: the architects of eukaryotic complexity, in EMBO Reports, vol. 2, nº 11, 2001, pp. 986–91, DOI:10.1093/embo-reports/kve230, PMC 1084129, PMID 11713189.
^ Mattick JS, Challenging the dogma: the hidden layer of non-protein-coding RNAs in complex organisms (PDF), in BioEssays : News and Reviews in Molecular, Cellular and Developmental Biology, vol. 25, nº 10, October 2003, pp. 930–9, DOI:10.1002/bies.10332, PMID 14505360 (archiviato dall'url originale il 6 marzo 2009).
^ Mattick JS, The hidden genetic program of complex organisms, in Scientific American, vol. 291, nº 4, October 2004, pp. 60–7, DOI:10.1038/scientificamerican1004-60, PMID 15487671 (archiviato dall'url originale l'8 febbraio 2015).
^ abc Wirta W, Mining the transcriptome – methods and applications, Stockholm, School of Biotechnology, Royal Institute of Technology, 2006, ISBN 91-7178-436-5, OCLC 185406288.
^ ab Rossi JJ, Ribozyme diagnostics comes of age, in Chemistry & Biology, vol. 11, nº 7, 2004, pp. 894–95, DOI:10.1016/j.chembiol.2004.07.002, PMID 15271347.
^ Gueneau de Novoa P, Williams KP e Williams, The tmRNA website: reductive evolution of tmRNA in plastids and other endosymbionts, in Nucleic Acids Res, vol. 32, Database issue, 2004, pp. D104–8, DOI:10.1093/nar/gkh102, PMC 308836, PMID 14681369.
^ RW Carthew e Sontheimer, EJ., Origins and Mechanisms of miRNAs and siRNAs., in Cell, DOI:10.1016/j.cell.2009.01.035, PMC 2675692, PMID 19239886. URL consultato il 20 febbraio 2009.
^ Kung-Hao Liang e Yeh, Chau-Ting, A gene expression restriction network mediated by sense and antisense Alu sequences located on protein-coding messenger RNAs., in BMC Genomics, DOI:10.1186/1471-2164-14-325, PMC 3655826, PMID 23663499. URL consultato l'11 maggio 2013.
^ Wu L e Belasco JG, Let me count the ways: mechanisms of gene regulation by miRNAs and siRNAs, in Mol. Cell, vol. 29, nº 1, January 2008, pp. 1–7, DOI:10.1016/j.molcel.2007.12.010, PMID 18206964.
^ Matzke MA e Matzke AJM, Planting the seeds of a new paradigm, in PLoS Biology, vol. 2, nº 5, 2004, pp. e133, DOI:10.1371/journal.pbio.0020133, PMC 406394, PMID 15138502.
^ Vazquez F, Vaucheret H, Rajagopalan R, Lepers C, Gasciolli V, Mallory AC, Hilbert J, Bartel DP e Crété P, Endogenous trans-acting siRNAs regulate the accumulation of Arabidopsis mRNAs, in Molecular Cell, vol. 16, nº 1, 2004, pp. 69–79, DOI:10.1016/j.molcel.2004.09.028, PMID 15469823.
^ Watanabe T, Totoki Y, Toyoda A, Kaneda M, Satomi Kuramochi-Miyagawa, Yayoi Obata, Hatsune Chiba, Yuji Kohara, Tomohiro Kono, Toru Nakano, M. Azim Surani, Y Sakaki e H Sasaki, Endogenous siRNAs from naturally formed dsRNAs regulate transcripts in mouse oocytes, in Nature, vol. 453, nº 7194, May 2008, pp. 539–43, Bibcode:2008Natur.453..539W, DOI:10.1038/nature06908, PMID 18404146.
^ Sontheimer EJ e Carthew RW, Silence from within: endogenous siRNAs and miRNAs, in Cell, vol. 122, nº 1, July 2005, pp. 9–12, DOI:10.1016/j.cell.2005.06.030, PMID 16009127.
^ Doran G, RNAi – Is one suffix sufficient?, in Journal of RNAi and Gene Silencing, vol. 3, nº 1, 2007, pp. 217–19 (archiviato dall'url originale il 16 luglio 2007).
^ Pushparaj PN, Aarthi JJ, Kumar SD e Manikandan J, RNAi and RNAa — The Yin and Yang of RNAome, in Bioinformation, vol. 2, nº 6, 2008, pp. 235–7, DOI:10.6026/97320630002235, PMC 2258431, PMID 18317570.
^ Horwich MD, Li C, Matranga C, Vagin V, Farley G, Wang P e Zamore PD, The Drosophila RNA methyltransferase, DmHen1, modifies germline piRNAs and single-stranded siRNAs in RISC, in Current Biology, vol. 17, nº 14, 2007, pp. 1265–72, DOI:10.1016/j.cub.2007.06.030, PMID 17604629.
^ Girard A, Sachidanandam R, Hannon GJ e Carmell MA, A germline-specific class of small RNAs binds mammalian Piwi proteins, in Nature, vol. 442, nº 7099, 2006, pp. 199–202, Bibcode:2006Natur.442..199G, DOI:10.1038/nature04917, PMID 16751776.
^ Horvath P e Barrangou R, CRISPR/Cas, the Immune System of Bacteria and Archaea, in Science, vol. 327, nº 5962, 2010, pp. 167–70, Bibcode:2010Sci...327..167H, DOI:10.1126/science.1179555, PMID 20056882.
^ Wagner EG, Altuvia S e Romby P, Antisense RNAs in bacteria and their genetic elements, in Adv Genet, Advances in Genetics, vol. 46, 2002, pp. 361–98, DOI:10.1016/S0065-2660(02)46013-0, ISBN 978-0-12-017646-5, PMID 11931231.
^ Gilbert SF, Developmental Biology, 7th, Sinauer, 2003, pp. 101–3, ISBN 0-87893-258-5, OCLC 154656422 154663147 174530692 177000492 177316159 51544170 54743254 59197768 61404850 66754122.
^ Amaral PP e Mattick JS, Noncoding RNA in development, in Mammalian genome : official journal of the International Mammalian Genome Society, vol. 19, 7–8, October 2008, pp. 454–92, DOI:10.1007/s00335-008-9136-7, PMID 18839252.
^ Heard E, Mongelard F, Arnaud D, Chureau C, Vourc'h C e Avner P, Human XIST yeast artificial chromosome transgenes show partial X inactivation center function in mouse embryonic stem cells, in Proc. Natl. Acad. Sci. USA, vol. 96, nº 12, 1999, pp. 6841–46, Bibcode:1999PNAS...96.6841H, DOI:10.1073/pnas.96.12.6841, PMC 22003, PMID 10359800.
^ Steitz TA e Steitz JA, A general two-metal-ion mechanism for catalytic RNA, in Proc. Natl. Acad. Sci. U.S.A., vol. 90, nº 14, 1993, pp. 6498–502, Bibcode:1993PNAS...90.6498S, DOI:10.1073/pnas.90.14.6498, PMC 46959, PMID 8341661.
^ Xie J, Zhang M, Zhou T, Hua X, Tang L e Wu W, Sno/scaRNAbase: a curated database for small nucleolar RNAs and cajal body-specific RNAs, in Nucleic Acids Res, vol. 35, Database issue, 2007, pp. D183–7, DOI:10.1093/nar/gkl873, PMC 1669756, PMID 17099227.
^ Omer AD, Ziesche S, Decatur WA, Fournier MJ e Dennis PP, RNA-modifying machines in archaea, in Molecular Microbiology, vol. 48, nº 3, 2003, pp. 617–29, DOI:10.1046/j.1365-2958.2003.03483.x, PMID 12694609.
^ Cavaillé J, Nicoloso M e Bachellerie JP, Targeted ribose methylation of RNA in vivo directed by tailored antisense RNA guides, in Nature, vol. 383, nº 6602, 1996, pp. 732–5, Bibcode:1996Natur.383..732C, DOI:10.1038/383732a0, PMID 8878486.
^ Kiss-László Z, Henry Y, Bachellerie JP, Caizergues-Ferrer M e Kiss T, Site-specific ribose methylation of preribosomal RNA: a novel function for small nucleolar RNAs, in Cell, vol. 85, nº 7, 1996, pp. 1077–88, DOI:10.1016/S0092-8674(00)81308-2, PMID 8674114.
^ Daròs JA, Elena SF e Flores R, Viroids: an Ariadne's thread into the RNA labyrinth, in EMBO Rep, vol. 7, nº 6, 2006, pp. 593–8, DOI:10.1038/sj.embor.7400706, PMC 1479586, PMID 16741503.
^ Kalendar R, Vicient CM, Peleg O, Anamthawat-Jonsson K, Bolshoy A e Schulman AH, Large retrotransposon derivatives: abundant, conserved but nonautonomous retroelements of barley and related genomes, in Genetics, vol. 166, nº 3, 2004, pp. 1437–50, DOI:10.1534/genetics.166.3.1437, PMC 1470764, PMID 15082561.
^ Podlevsky JD, Bley CJ, Omana RV, Qi X e Chen JJ, The telomerase database, in Nucleic Acids Res, vol. 36, Database issue, 2008, pp. D339–43, DOI:10.1093/nar/gkm700, PMC 2238860, PMID 18073191.
^ T. Blevins, R. Rajeswaran, PV. Shivaprasad, D. Beknazariants, A. Si-Ammour, HS. Park, F. Vazquez, D. Robertson e F. Meins, Four plant Dicers mediate viral small RNA biogenesis and DNA virus induced silencing, in Nucleic Acids Res, vol. 34, nº 21, 2006, pp. 6233–46, DOI:10.1093/nar/gkl886, PMC 1669714, PMID 17090584.
^ Jana S, Chakraborty C, Nandi S e Deb JK, RNA interference: potential therapeutic targets, in Appl. Microbiol. Biotechnol., vol. 65, nº 6, 2004, pp. 649–57, DOI:10.1007/s00253-004-1732-1, PMID 15372214.
^ Schultz U, Kaspers B e Staeheli P, The interferon system of non-mammalian vertebrates, in Dev. Comp. Immunol., vol. 28, nº 5, 2004, pp. 499–508, DOI:10.1016/j.dci.2003.09.009, PMID 15062646.
^ K. A. Whitehead, J. E. Dahlman, R. S. Langer e D. G. Anderson, Silencing or Stimulation? SiRNA Delivery and the Immune System, in Annual Review of Chemical and Biomolecular Engineering, vol. 2, 2011, pp. 77–96, DOI:10.1146/annurev-chembioeng-061010-114133, PMID 22432611.
^ Dahm R, Friedrich Miescher and the discovery of DNA, in Developmental Biology, vol. 278, nº 2, 2005, pp. 274–88, DOI:10.1016/j.ydbio.2004.11.028, PMID 15680349.
^ Caspersson T e Schultz J, Pentose nucleotides in the cytoplasm of growing tissues, in Nature, vol. 143, nº 3623, 1939, pp. 602–3, Bibcode:1939Natur.143..602C, DOI:10.1038/143602c0.
^ Ochoa S, Enzymatic synthesis of ribonucleic acid (PDF), su Nobel Lecture, 1959.
^ Rich A e Davies, D, A New Two-Stranded Helical Structure: Polyadenylic Acid and Polyuridylic Acid, in Journal of the American Chemical Society, vol. 78, nº 14, 1956, pp. 3548–3549, DOI:10.1021/ja01595a086.
^ Holley RW, J. Apgar, G. A. Everett, J. T. Madison, M. Marquisee, S. H. Merrill, J. R. Penswick e A. Zamir, Structure of a ribonucleic acid, in Science, vol. 147, nº 3664, 1965, pp. 1462–65, Bibcode:1965Sci...147.1462H, DOI:10.1126/science.147.3664.1462, PMID 14263761.
^ Siebert S, Common sequence structure properties and stable regions in RNA secondary structures (PDF), su Dissertation, Albert-Ludwigs-Universität, Freiburg im Breisgau, 2006, p. 1 (archiviato dall'url originale il 9 marzo 2012).
^ Szathmáry E, The origin of the genetic code: amino acids as cofactors in an RNA world, in Trends Genet, vol. 15, nº 6, 1999, pp. 223–9, DOI:10.1016/S0168-9525(99)01730-8, PMID 10354582.
^ Fiers W, R. Ysebaert, F. Duerinck, G. Haegeman, D. Iserentant, J. Merregaert, W. Min Jou, F. Molemans, A. Raeymaekers, A. Van Den Berghe, G. Volckaert e M Ysebaert, Complete nucleotide-sequence of bacteriophage MS2-RNA: primary and secondary structure of replicase gene, in Nature, vol. 260, nº 5551, 1976, pp. 500–7, Bibcode:1976Natur.260..500F, DOI:10.1038/260500a0, PMID 1264203.
^ Napoli C, Lemieux C e Jorgensen R, Introduction of a chimeric chalcone synthase gene into petunia results in reversible co-suppression of homologous genes in trans, in Plant Cell, vol. 2, nº 4, 1990, pp. 279–89, DOI:10.1105/tpc.2.4.279, PMC 159885, PMID 12354959.
^ Dafny-Yelin M, Chung SM, Frankman EL e Tzfira T, pSAT RNA interference vectors: a modular series for multiple gene down-regulation in plants, in Plant Physiol., vol. 145, nº 4, December 2007, pp. 1272–81, DOI:10.1104/pp.107.106062, PMC 2151715, PMID 17766396.
^ Ruvkun G, Glimpses of a tiny RNA world, in Science, vol. 294, nº 5543, 2001, pp. 797–99, DOI:10.1126/science.1066315, PMID 11679654.
^ Fichou Y e Férec C, The potential of oligonucleotides for therapeutic applications, in Trends in Biotechnology, vol. 24, nº 12, 2006, pp. 563–70, DOI:10.1016/j.tibtech.2006.10.003, PMID 17045686.
Bibliografia |
- David L. Nelson, Michael M. Cox, I Principi di Biochimica di Lehninger, 3ª ed., Bologna, Zanichelli, febbraio 2002, ISBN 88-08-09035-3.
- (EN) R.N. Shukla, Analysis of Chromosomes, ISBN 978-93-84568-17-7.
- (EN) Berg JM, Tymoczko JL e Stryer L, Biochemistry, 5th, WH Freeman and Company, 2002, ISBN 0-7167-4684-0, OCLC 179705944 48055706 59502128.
- (EN) Barciszewski J, Frederic B e Clark C, RNA biochemistry and biotechnology, Springer, 1999, ISBN 0-7923-5862-7, OCLC 52403776.
- (EN) Jankowski JAZ e Polak JM, Clinical gene analysis and manipulation: Tools, techniques and troubleshooting, Cambridge University Press, 1996, ISBN 0-521-47896-0, OCLC 33838261.
- (EN) Söll D e RajBhandary U, TRNA: Structure, biosynthesis, and function, ASM Press, 1995, ISBN 1-55581-073-X, OCLC 183036381 30663724.
- (EN) Cooper GC e Hausman RE, The Cell: A Molecular Approach, 3rd, Sinauer, 2004, ISBN 0-87893-214-3, OCLC 174924833 52121379 52359301 56050609.
- (EN) Gilbert SF, Developmental Biology, 7th, Sinauer, 2003, ISBN 0-87893-258-5, OCLC 154656422 154663147 174530692 177000492 177316159 51544170 54743254 59197768 61404850 66754122.
- (EN) Wirta W, Mining the transcriptome – methods and applications, Stockholm, School of Biotechnology, Royal Institute of Technology, 2006, ISBN 91-7178-436-5, OCLC 185406288.
Voci correlate |
- Complementarità
- DNA
Altri progetti |
Altri progetti
- Wikimedia Commons
 Wikimedia Commons contiene immagini o altri file su RNA
Wikimedia Commons contiene immagini o altri file su RNA
Collegamenti esterni |
RNA, su thes.bncf.firenze.sbn.it, Biblioteca Nazionale Centrale di Firenze.
- (EN) IUPAC Gold Book, "ribonucleic acids (RNA)", su goldbook.iupac.org.
- L'RNA ribosomico, sul portale RAI Scuola, su raiscuola.rai.it.
- [1]
.mw-parser-output .navbox{border:1px solid #aaa;clear:both;margin:auto;padding:2px;width:100%}.mw-parser-output .navbox th{padding-left:1em;padding-right:1em;text-align:center}.mw-parser-output .navbox>tbody>tr:first-child>th{background:#ccf;font-size:90%;width:100%}.mw-parser-output .navbox_navbar{float:left;margin:0;padding:0 10px 0 0;text-align:left;width:6em}.mw-parser-output .navbox_title{font-size:110%}.mw-parser-output .navbox_abovebelow{background:#ddf;font-size:90%;font-weight:normal}.mw-parser-output .navbox_group{background:#ddf;font-size:90%;padding:0 10px;white-space:nowrap}.mw-parser-output .navbox_list{font-size:90%;width:100%}.mw-parser-output .navbox_odd{background:#fdfdfd}.mw-parser-output .navbox_even{background:#f7f7f7}.mw-parser-output .navbox_center{text-align:center}.mw-parser-output .navbox .navbox_image{padding-left:7px;vertical-align:middle;width:0}.mw-parser-output .navbox+.navbox{margin-top:-1px}.mw-parser-output .navbox .mw-collapsible-toggle{font-weight:normal;text-align:right;width:7em}.mw-parser-output .subnavbox{margin:-3px;width:100%}.mw-parser-output .subnavbox_group{background:#ddf;padding:0 10px}


.mw-parser-output .CdA{border:1px solid #aaa;width:100%;margin:auto;font-size:90%;padding:2px}.mw-parser-output .CdA th{background-color:#ddddff;font-weight:bold;width:20%}
| Controllo di autorità | LCCN (EN) sh85113850 · GND (DE) 4076759-0 · BNF (FR) cb12175223g (data) |
|---|


